Dureux, A., Zanini, A., Menon, R. S., & Everling, S. (2024). Vocal processing networks in the human and marmoset brain. Neuroscience. https://doi.org/10.1101/2024.09.15.613152
Dureux (2024)
Abstract
Understanding the brain circuitry involved in vocal processing across species is crucial for unraveling the evolutionary roots of human communication. While previous research has pinpointed voice-sensitive regions in primates, direct cross-species comparisons using standardized protocols are limited. This study utilizes ultra-high field fMRI to explore vocal processing mechanisms in humans and marmosets. By employing voice-sensitive regions of interest (ROIs) identified via auditory localizers, we analyzed response time courses to species- specific vocalizations and non-vocal sounds using a dynamic auditory-stimulation paradigm. This approach gradually introduced sounds into white noise over 33 seconds. Results revealed that both species have responsive areas in the temporal, frontal, and cingulate cortices, with a distinct preference for vocalizations. Significant differences were found in the response time courses between vocal and non-vocal sounds, with humans displaying faster responses to vocalizations than marmosets. We also identified a shared antero-ventral auditory pathway in both species for vocal processing, originating from the superior temporal gyrus. Conversely, a posterior-dorsal pathway was more prominent in humans, whereas in marmosets, this pathway processed both sound types similarly. This comparative study sheds light on both conserved and divergent auditory pathways in primates, providing new insights into conspecific vocalization processing.
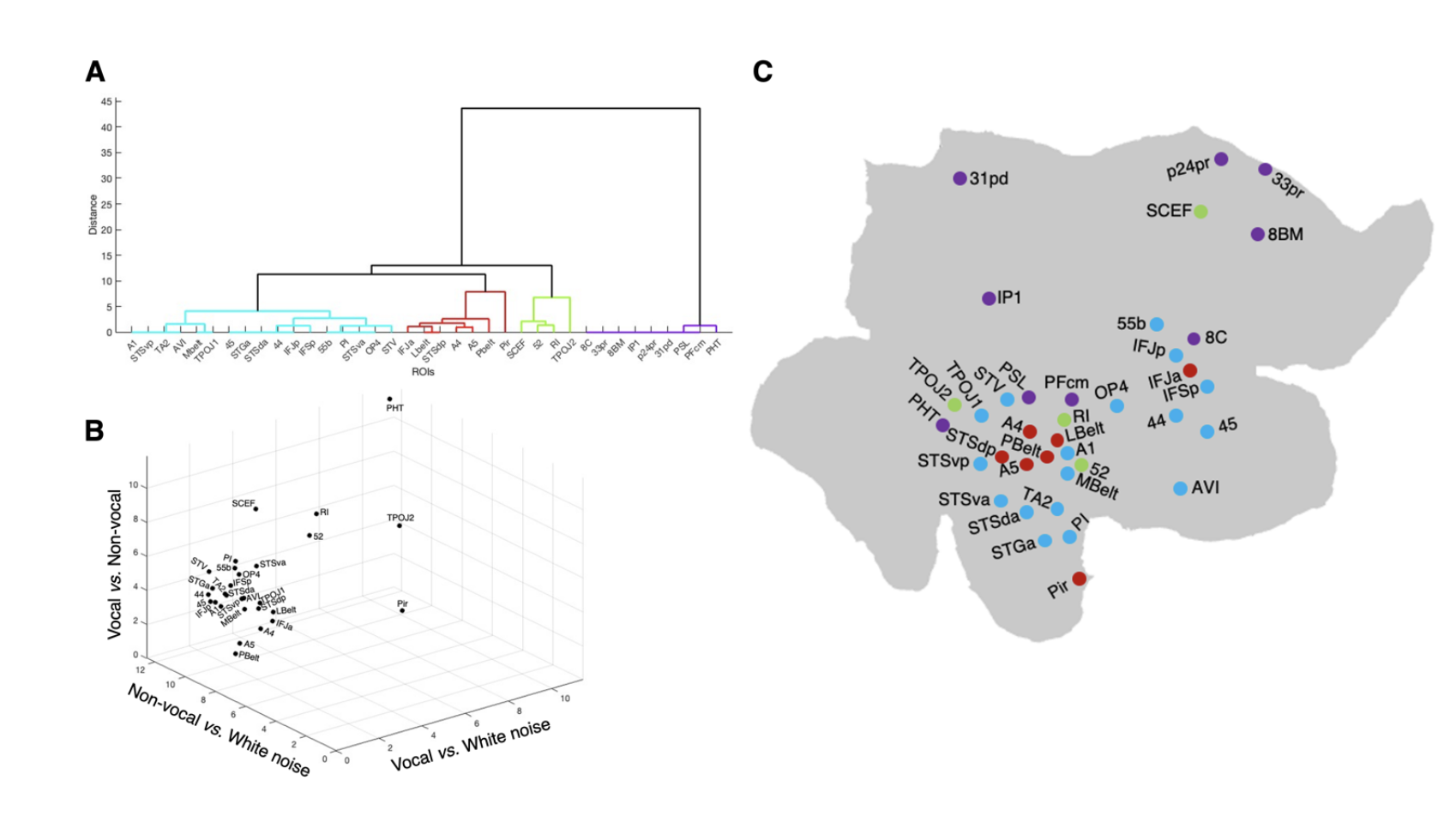
In humans (Figure 2), activations predominantly occurred within the superior temporal gyrus and adjacent insular cortex across all contrasts, including early auditory areas such as A1, 52, and the MedialBelt (MBelt), LateralBelt (LBelt), and ParaBelt (PBelt) Complexes. Additional activations included the retroinsular cortex (RI), para-insular area (PI), and Pir area, alongside auditory association areas A4, A5, and TA2. Posterior temporal cortex activations encompassed areas PFcm, periSylvian language area (PSL), temporo-parieto-occipital TPOJ1 and TPOJ2, PHT, and STV, as well as auditory association areas STSd posterior (STSdp), and STSv posterior (STSvp). The anterior temporal cortex showed activations in STSv anterior (STSva) and STSd anterior (STSda) areas. Additional activations were also present in lateral prefrontal and inferior frontal areas 55b, 8C, IFJp, IFJa, IFSp, 44, 45, as well as the posterior opercular area (OP4) and the anterior ventral insular area (AVI). Some medial prefrontal and cingulate areas, predominantly in the right hemisphere, were also activated, including 8BM, SCEF, p24pr, 33pr, and 31pd.
Several regions, including A1, LBelt, PBelt, MBelt, A4, A5, area 52, TPOJ1, IFSp, IFJa, IFJp, and STSdp, displayed increasing responses to both vocalizations and non-vocal sounds, with notably earlier and more pronounced responses to vocalizations, suggesting heightened sensitivity to these socially relevant sounds. Notably, areas such as PI, TA2, OP4, 45, 44, 55b, SCEF, STGa, and STSda responded almost exclusively to vocalizations, showing minimal or no response to non vocal sounds and pure white noise.
Conversely, RI, Pir, TPOJ2, PHT, STV, AVI, and STSva, demonstrated very weak responses to vocalizations. Additional areas, including PSL, PFcm, IP1, and medial and lateral prefrontal areas 8C, 31pd, p24pr, 33pr, and 8BM, remained unresponsive to all tested stimuli.
a cluster in purple included regions such as 8C, 33pr, 8BM, IP1, p24pr, 31pd, PSL, PFcm, and PHT, showing no responses to auditory stimuli (Figure 8A and Figure 8C). These regions were positioned away from the main axes in the 3D plot, indicating minimal involvement in the dynamic auditory-stimulation paradigm (Figure 8B)
A prominent cluster, depicted in light blue, encompassed regions that significantly distinguished between vocalizations and both white noise and non-vocal sounds. However, these regions did not differentiate between non-vocal sounds and white noise, or they only did so when the non-vocal sound presence reached 100%. This cluster included human prefrontal areas 44, 45, IFJp, IFSp, OP4, and the premotor area 55b, along with areas in the superior temporal gyrus such as early auditory areas A1, 52, RI, and auditory association areas TA2 and STGa. It also comprised anterior temporal areas STSda, STSva, posterior temporal areas STSvp, STV, adjacent insular areas AVI, PI, and cingulate area SCEF, alongside marmoset cingulate areas 32 and 32v, and the anterior and ventral temporal area Pir.
nice to know:
- human auditory regions including A4, A5, LBelt, PBelt, STSdp, and the inferior frontal area IFJa responded to non-vocal sounds much later, typically between 70-90%, with some superior temporal gyrus areas (A1, TA2, 52, MBelt) and posterior temporal areas (TPOJ1, STSvp) only responding at 90-100%.
see also
Tags: neuroscience science
Superlink: 050 🧠Neuroscience
Source
Created: 2025-12-02 15:04